| Home |
|
|
|
|
Figure 6-1: possible distribution of
dorsal and
ventral hair colour zones; names of facial markings and some
pigmented
skin areas
Figure 6-2: examples for facial and
dorsal
fur colour patterns occurring in lorises and pottos
Propositions for standardized determination
and
documentation of coat colour and colour patterns on trunk and
limbs
References
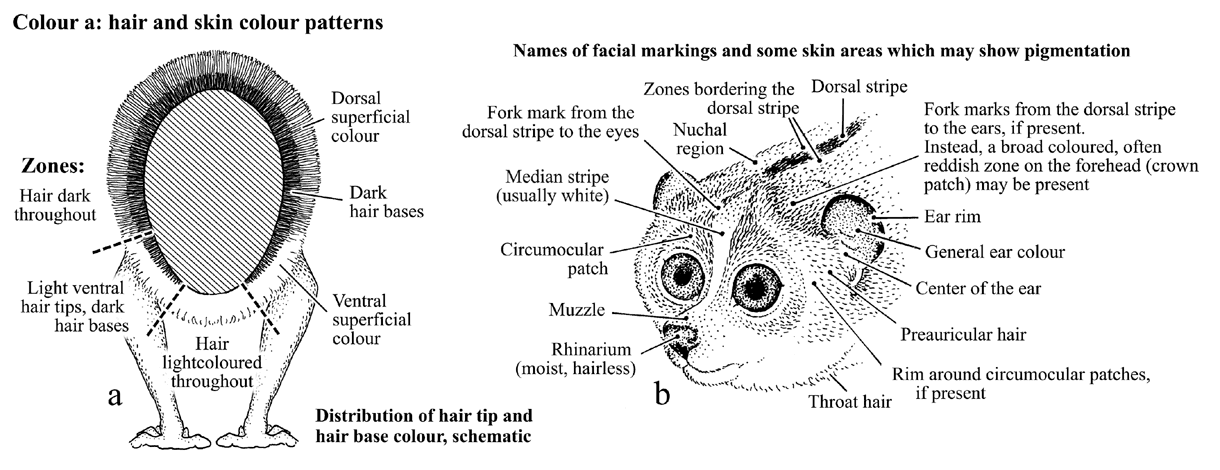
Figure 6-1: possible distribution of dorsal and ventral hair colour zones; names of facial markings and some pigmented skin areas
a: Distribution of dorsal and ventral colour zones:
In some mammal species, the extent of dorsal, ventral superficial
fur
colour, hair base colour and their changes during seasonal molt
may be
of considerable importance for taxonomic identification (Sumner
1926, 1927).
The figure proposes a way to locate the borders of distinct zones.
b: Names and location of possible facial markings and names of
some
parts of the face which may show skin pigmentation
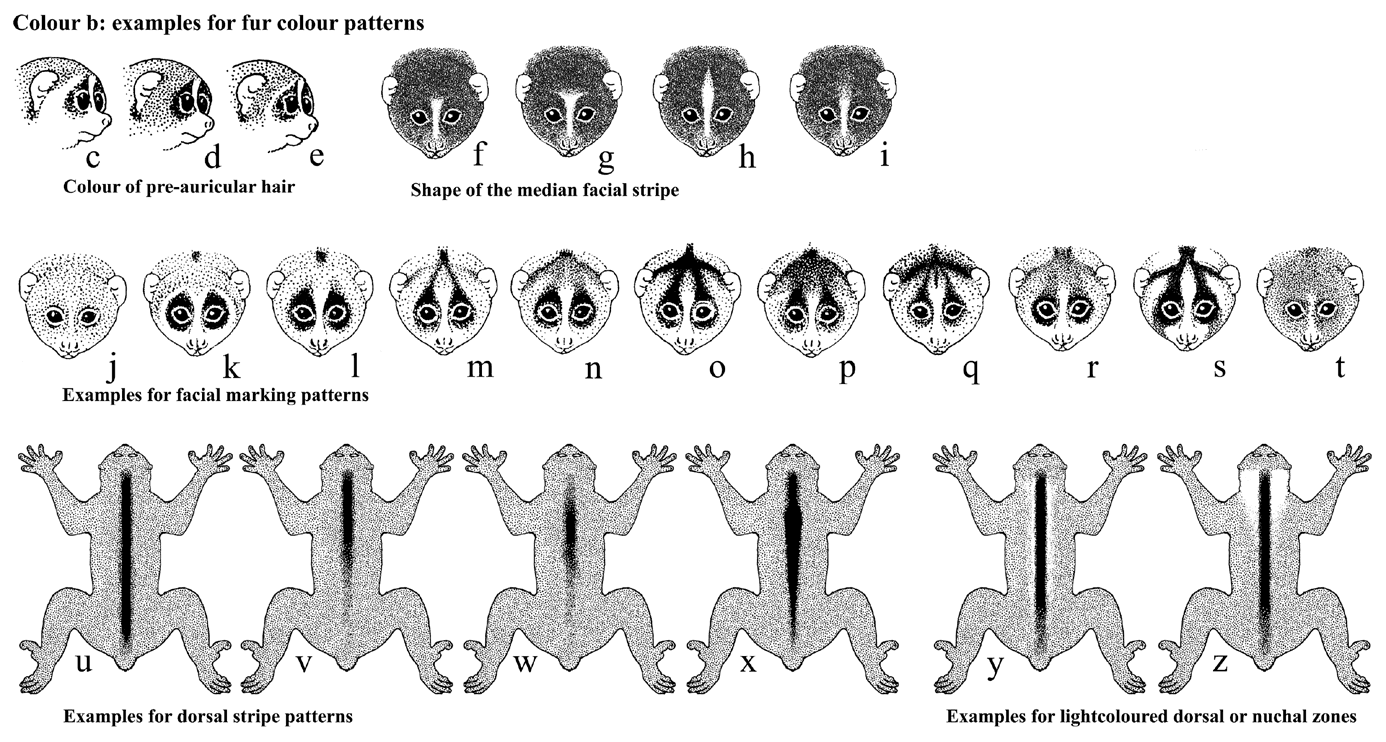
Figure 6-2: examples
for facial
and dorsal fur colour patterns occurring in lorises and pottos
The examples in the following review in figure 6-2 may be used to
record
external features of lorises by noting the number of the
example(s) most
similar
to the feature seen in the examined animal. (See also in this
chapter:
drafts for measuring and description form sheets for taxonomic
purposes)
Colour of preauricular hair:
c: White or very light-coloured. d: Grey or
otherwise
coloured, not whitish. e: white zone between darker
preauricular
hair and dark circumocular patches (typical for L. t. grandis,
according
to Osman Hill, 1953).
f-i: possible shape of the light-coloured median facial
stripe:
f: Stripe ending rather abruptly above eye-level, dorsal
tip
not pointed. g: Ending above-eye-level, bifurcating
above.
h:
Pointed dorsally, reaching up or almost upo to the dorsal stripe
(the upper
end may be slightly darker). i: Extending well
above
eye-level. but fading dorally, no clear upper end.
j-t: Facial marking patterns occurring in lorises and pottos:
j: Light-coloured face with no evident dark or reddish
markings;
a darker dorsal stripe and a lighter-coloured to white median
stripe may
be present. k: Dark circumocular patches present,
surrounded by
a light-coloured zone, white median stripe bifurcating above as in
fig.
g, no darker zones or stripes connecting the circumocular patches
with
the dorsal stripe, if the latter is present . l: Dark
circumocular
patches present, no darker zones or stripes connecting the
circumocular
patches with the dorsal stripe, if the latter is present, no
lighter zone
around the circumocular patches. (Said to be typical for L.
t./l. grandis;
also occurring in some nordicus). m: Darker
stripes
(usually faint) connecting the circumocular patches and dorsal
stripe.
Darker zones / stripes between dorsal stripe and ears either
absent ot
faint and incomplete. (Occurring for instance in southern N.
bengalensis,
old synonym N. c. tenasserimensis) n:
Dark stripes
connecting the ears with the dorsal stripe. If other facial stripe
patterns
or zones are present, they are lighter-coloured and indistinct.
(According
to Osman Hill occurring in Bornean slow lorises). o:
Well-visible
dark stripes connecting both circumocular patches and ears with
the dorsal
stripe. Median lighter stripe as in figure f or i. p:
Instead
of a dark forkmark (stripe) pattern, a broader zone or crown
patch, usually
reddish, extends over the forehead, up to the ears and
circumocular patches.
(Occurring in N. c. coucang). q: Facial
stripe pattern
as in figs o or p, but in addition the upper part of the median
white stripe
includes a median dark little stripe. (Occurring in some N.
pygmaeus).
r:
Forkmark (stripe) pattern present, but indistinct. (Occurring in N.
c.
menagensis) s: Clear-marked dark (usually
blackish,
sometimes brown or grey) stripes on a light-coloured, whitish
face. Median
stripe as in fig. h, reaching up to the dorsal stripe, dividing
the dark
pattern into two branches which divide again to form stripes to
ears and
eyes. From the circumocular patches, usually a dark zone extends
further
down to the corners of the mouth. (Typical Javan loris face). t:
Face of uniform reddish, brown or dark colour; a median light
stripe, lighter
cheeks and / or a dark dorsal stripe may be present or absent; no
dark
stripes leading to ears or eyes (Occurring in the African pottos).
Dorsal colour patterns:
u-x: Shape, extent of the dorsal stripe:
u: Dark or reddish stripe extending over the entire baxk
(the
caudal end may be slightly fainter, but is still well
visible. v:
Dark, reddish or red stripe in the thoracal region; caudally faint
or absent.
w:
Dorsal stripe faint, indistinct; only a short part in the thoracal
region
distinct, dark. x: Stripe broadest in the shoulder
region,
becoming narrower caudally rather than fading.
Light-coloured zones in the dorsal region:
y: A narrow lighter-coloured zone may be present along the
sides
of the stripe. z: Zone in the nuchal region more
light-coloured
than the surrounding dorsal fur. This area may be only slightly
lighter
than the back (frequent in all loris forms) or light creamy to
whisish,
strongly contrasting with back and dorsal stripe (the latter is
typical
for Javan lorises). The light zone may extend caudally as in
fig.
yalong the dorsal stripe as lighter colour or frosting, gradually
fading
caudally.
In addition to the examples shown here, the dorsal stripe may be clear-cut or blunt, broad or narrow, of uniform colour or for instance brown or reddish with darker rims. In some museum skins it looks wavy or zigzags; this might be a consequence of preservation. In animals with a light-coloured nuchal zone as shown in fig. z, the part of the dorsal stripe crossing this zone may also be lighter-coloured than the more caudal parts of the stripe.
Propositions for
standardized
determination and documentation of coat colour and colour
patterns on trunk
and limbs
Usually in literature colours are described with words. A
dictinary
of colour terms and some more information about colour standards
are for
instance available in the website http://www.anthus.com/Colors/NBS.html
by David Mundie. Reproducible documentation and description of
pelage colours
would require a standard light quality (usually a spectral
composition
resembling midday daylight) and some sort of generally available
colour
standard or a colorimetric apparatus. Portable spektrometers and
accessories
can for instance be obtained from: Ocean Optics Europe, Soerense
Zand 4,
NL-6961 LL Eerbeek, The Netherlands, phone: x31 / (0)313 670 170,
FAX:
x31 / (0)313 670 179.
A source of standard light should be easily available; halogen light and usual photo flashlight are rather similar to daylight. Accumulator / battery headlamps for field work, allowing a choice between bright halogen light and more dim "standard" light for longer observation, are for instance available from Petzl Distribution Sport: Zone Industrielle, 38920 Crolles, France. Phone: x33-476 92 09 20; FAX: x33-476 08 82 04; petzl@dial.oleane.com; http:\\www.petzl.com.
Colour determination: Colorimetric measuring of spectral composition of the light reflected by the fur would be the most exact method to determine fur colour. The colorimetric method Sumner (1927) described, however, requires killing and preparation of specimens and an apparatus which is usually not available in the field. Comparison of fur colour of live specimens with printed colour tables with the human eye is not quite exact, but seems to be the method which can be applied best in the field. Printed standard colours on paper may fade after some time; so standard colour tables should not be too old.
The structure of fur with shadows and gleaming hair tips make a comparison with the homogenus surface of coloured paper difficult, and light from cranial or caudal direction produces different appearance of colours. Sumner used a light direction perpendicular to the direction of hair and pressed down the hair with a cover glass. Küppers (1981) recommends to cover adjacent areas with grey paper or cardboard leaving only the colour in question visible because neighbouring colours influence the subjective perception of a colour. A colour table which tries to combine these recommendations is in preparation.
In addition, mammalian pelage, even in areas of relatively uniform-looking colour, usually is not homogenous, but consists of a mixture of differently-coloured types of hair and of hairs showing a succession of different colours from base to tip. In mice, Sumner (1926, 1927) distinguished three types of coat colour zones: one (dorsally) in which hairs are coloured throughout, one (ventrally) in which the entire hair is devoid of pigment and white, and a transition zone between both in which hairs are pigmented at the base, but lacking pigment more distally. Sumner regarded the relative extent of these colour zones as a racial character of considerable importance in certain mouse species. Similar patterns occur in Loris, and Osman Hill (1933, 1953) regarded them as a help for instance for distinguishing Loris tardigradus lydekkerianus (with ventral hair white throughout) from the similar L. t. nordicus (with dark ventral hair bases); at Ruhr-University, however, some L. t. nordicus specimens had a narrow median zone of entirely white hair.
In lorises annd pottos, colour patterns of the fur may include
certain
markings (dark stripes, circumocular patches, other). Frosting
(white tips
of dark dorsal hair) may be present or absent. A dark dorsal
stripe may
be present, it is usually most distinct in the thoracic region of
the trunk
and may, in some specimens, be enclosed by light-coloured stripes
on either
side or a broader light-coloured zone for instance extending over
the entire
nuchal region.
Natural variation of coat colours (adult specimens) within
populations
/ species
Occurrence of different colour types (red and grey varieties,
random
erythrism) in one mammal species is known (Sanderson, 1940). Osman
Hill
and Phillips (1932) have already mentioned that for instance all
slender
loris forms "present wide individual variation" in coat colour,
with additional
age and sex variations, and that only the average coloration of
several
specimens of a population in connection with other differences may
serve
to distinguish them from other forms. In L. t. nordicus
from Polonnaruwa,
adult specimens of dark grey to light yellowish colour with strong
to almost
absent frosting occur within one natural breeding population (data
from
Ruhr-University). In Nycticebus coucang coucang observed
in Selangor,
Malaysia, too, a considerable variation of coat colour was found
(Wiens,
Zitzmann, pers. comm.).
Age-related variation of coat colour
The coat colour of infants and juveniles usually differs from that
of adults. In L. t. nordicus, for example, juveniles show
a light
yellowish to reddish fur without frosting (white hair tips), with
reddish
circumocular patches; they might be mistaken for specimens of the
smaller
reddish forms by observers not familiar with lorises. Adult colour
of L.
t. nordicus, usually grey, seldom yellowish-grey,
increasingly develops
up to the age of about 16 to 21 month. In ageing animals, the fur
of the
face, and to a lesser degree of other parts, may become
increasingly grey
to almost white. Development of white hair may start at the age of
5 years
in some specimens, others show no white hair at the age of 13
years or
more.
External / artificial causes of variation of coat colours;
influence
of preservation methods: see also field
methods and necropsy chapter, part about colour
changes
by preservation
In addition to inherited and age-related variation of fur colour,
the
colour of preserved or otherwise treated specimens may differ from
natural
colour of the animals. Sumner (1927) who cleaned fur with benzine
or other
chemical agents for better comparability mentions colour changes.
According
to Sanderson (1940), changes of colour of hair in museum
specimens
may occur due to fading because of old age or exposure to light,
due to
drying by smoke, proximity of certain chemicals, radiators and
other influences.
Series of skins dried in bright sun in the field tended to turn
reddish.
Experiments with a maroon-coloured rat, Malacomys longipes,
showed
that furs became grey, dark brown or reddish-brown, according to
whether
they were dried in a closed container, in shade or bright
sunlight. Series
of
Praomys, including examples of bright reddish and
olive-grey
varieties, could be evenly dried to a corresponding dull siena
when submitted
to reciprocal treatment. Some reddish hair may be dyed by plant
juice.
Greenish coloration in certain arboreal species may be due to
algae as
in the sloth Bradypus; a bright green dorsal colour in Galagoides
demidoff, which fades rapidly after the death of the animal
and turns
rusty, might also be caused by algae (Sanderson 1940).
Küppers, H., 1981: DuMont´s Farben-Atlas. DuMont Buchverlag, Köln.
Osman Hill, W. C.; Phillips, W. W. A., 1932: A new race of slender loris from the Highlands of Ceylon. Ceylon Journal of Science / Spolia Zeylanica XVII (2): 109-123.
Sanderson, I. T., 1940: The mammals of the north Cameroon forest area. Transactions of the Zoological Society of London 14: 623-725
Ridgway, R., 1912: Color standards and color nomenclature. Published by the author. Washington. 44 pp., 53 coloured plates.
Sumner, F. B., 1926: An analysis of geographic variation in mice of the Peromyscus polionotus group from Florida and Alabama. Journal of Mammalogy 7: 149-184 (+ 4 plates).
Sumner, F. B., 1927: Linear and colorimetric measurements
of
small mammals. Journal of Mammalogy 8 (3): 177-206.
Some literature concerning colour standards:
Grum, F.; Bartleson, C. J. (eds.), 1980: Optical radiation measurements volume 2: color measurement. Academic Press, New York, London [u. a.].
Jacobs, G. H., 1981: Comparative color vision. Academic Press, New York, London [u. a.].
MacAdam, D. L., 1985: Color measurement. Theme and variations. Springer Verlag, Berlin [u. a.].
Munsell Color Co., 1971: Munsell soil color charts. 13 tables, 8 pp.. Baltimore, MD.
Munsell Color Co., 1972: Munsell Book of Color. Glossy Finish Collection. Removable samples in two binders, 1966, 2.5 BG-10 RP. 1-2. Baltimore, MD.
Ridgway, R., 1912: Color standards and color nomenclature. Published by the author. Washington. 44 pp., 53 coloured plates.
Smithe, F. B., 1975-1981: Naturalist´s color guide.
American
Museum of Natural History, New York, several parts.
|
Lorises and pottos: species,
subspecies, local
populations. In: http://www.species.net
|
Last amendment: 20 March 2003
|