Survey and field research methods: detection and identification of
lorises
and pottos in the wild, some literature about methods and
sympatric species
Compiled by Helga Schulze with
data
by (in alphabetic order) Prof. Dr. Eberhard Curio, Bernhard Meier,
Anna
Nekaris, Ulrike Streicher (Endangered Primate Rescue Center),
Frank Wiens
and Annette Zitzmann.
Loris tardigradus
(individual differences
see identification key for subspecies)
Nycticebus sp.
Paradoxurus hermaphroditus,
common palm civet
Felis chaus, jungle cat
Martes sp., marten
Herpestes sp.,
mongooses (basically terrestrial)
Some African species:
Arctocebus and Perodicticus
Galagos:
.
Euoticus sp.
Galagoides demidoff
In darkness
with a torchlight, head lamp or other light. When the
source of light is
held close to the eyes of the observer, the tapetum
lucidum in the
eyes of the animals (reflecting layer of cells behind
the retina) causes
a reflection well visible as long as the animal is
looking towards the
light. The tapetum causes light rays to cross the retina
twice and probably
shifts ultra-violet light into visible wavelength for
better vision in
darkness. It occurs in many nocturnal and some diurnal
and crepuscular
species (see below); differences in the colour and
strength of the eye-shine,
the size and distance of eyes provide some information
about identity of
species. Even light of rather low intensity causes a
visible eye-shine
in Loris; the bright reflexion of N. coucang
eyes is well
visible from a distance of 200 m in the wild (B. Meier,
unpublished; Petter,
Hladik 1970; Barrett, 1984; F. Wiens and A. Zitzmann,
pers. comm.).
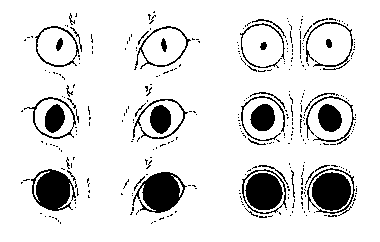
Change of pupil size and shape with diminishing
light intensity.
Left: cat. Right: Loris
Sources
of
light used or recommended:
Use of red or infrared light
(see
below, technical equipment) has a number of advantages: it is
not strongly
reflected by surrounding vegetation, and at least some nocturnal
animal
species are unable or less able to see these wavelengths.
Barrett (1984)
confirmed this effect for Nycticebus. The animals are
therefore
less aware of observation, show a more normal behaviour and a
diminished
tendency to retreat or hide their faces for camouflage.
Disadvantages of
red light: it does not allow proper judgement of the colour of
eyeshine,
and its diminished intensity may be still sufficient in more
open vegetation;
in tropical rainforest, loss of light by canopy may make use of
the stronger
white light necessary (Southern 1955; Barrett 1984).
Lamps used by several
authors: Barrett
(1984): headlamp powered by a 6 v motor-cycle accumulator
battery which
delivered a steady current for 24-30 hours. Charles-Dominique
(1971): strong
white head-lamp with a light angle wider than the field of
vision and two
nickel cadmium accumulators fixed to a girdle. B. Meier
(unpublished):
strong pivoted spot-light (white) on top of a Landrover.
Southern (1955) tried
infrared light
and visible red light for observation; since military infrared
"sniperscopes"
provided an insufficient resolution and the high-pitched whine
emitted
by the transformer disturbed the animals, he preferred use of
normal binoculars
in combination with an automobile headlamp screened like a
darkroom lamp
to give visible red light.
Headlamps can for instance be
obtained
from the following sources:
Petzl Distribution Sport:
Zone Industrielle,
38920 Crolles, France. Phone: x33-476 92 09 20; FAX: x33-476 08
82 04;
petzl@dial.oleane.com; http://www.petzl.com:
accumulator / battery headlamps including one which allows
choice between
bright halogen light with shorter duration and more dim
"standard" light
for longer observation with one set of batteries. This type of
lamp, when
equipped with an additional red filter in front of one lamp,
might provide
further possibilities to observe lorises.
CEAG Licht- und
Stromversorgungstechnik,
Postfach 305, Dortmund, Germany (accumulator lamps, including
strong headlamps,
for minery and other purposes)
Characteristics of eye
reflection
and other features helpful for detection and identification:
A tapetum is present in all
prosimians
with the exception of few diurnal and crepuscular Lemur
species,
but absent in tarsiers and in simian primates; Aotus
lacks an eyeshine
in spite of being nocturnal and having a kind of tapetum (Martin
1990,
Rowe 1996). A conspicuous eyeshine is, for instance, also found
in many
nocturnal carnivores, particularly in cats, in some marsupials,
rodents
and fruit-bats (e. g. Pteropus) and in some birds
(Boeckh, 1975;
Grzimek, 1967; Martin, 1990)
Eyeshine characteristics:
In prosimian primates in
general:
golden-yellow or yellow-red (Martin, 1990)
In Loris: eye
reflexion
strong; when the eyes of the animal are directed towards the
source of
light, two large round orange spots, set very close to each
other (see
figure above), become visible. According to Singh et al. (2000)
eye reflection
in L. t. lydekkerianus in the fiels was orange from any
direction whereas
in other nocturnal mammals, for instance civets, eye colour
varied with
eye movements; civet eyes in addition were smaller. Typical way
to glance
for a moment at intervals, see also below: behavioural
characteristics.
Eye characteristics cannot be mistaken for those of sympatric
other species
in Sri Lanka; no such statement for India found. (B. Meier,
pers. comm.;
Petter, Hladik 1970; Nekaris 1997)
In Nycticebus pygmaeus:
as in N. coucang
In Nycticebus coucang:
strong orange or orange-red reflexion (Barrett 1984; F. Wiens,
pers. comm.);
because of similarity with other sympatric species (see below)
characteristics
of eye-shine alone are not sufficient for species
identification, consideration
of characteristic movement and eye-blinking patterns may be
helpful (F.
Wiens, pers. comm.)
In Arctocebus:
?
In Perodicticus:
?
Examples of sympatric
species
in Asia which might be mistaken for lorises because of
eye-shine:
Carnivores: in general:
eyeshine
greenish-blue (Martin, 1990) or yellow (see below). Arboreal
nocturnal
carnivores in Sri Lanka: yellow-green reflexion (B. Meier, pers.
comm.;
Petter, Hladik 1970). Viverrids in Malaysia: some species with
an eye-shine
similar to that of N. coucang, but different movements
and eye-blinking
patterns (F. Wiens, pers. comm.). Palm civets, particularly Paradoxurus
hermaphroditus, in Malaysia: yellow to orange, somewhat
similar to
N.
coucang (F. Wiens, pers. comm.). (Barrett, 1984, mentions
white or
lemon-yellow reflexion). Cats: yellowish eyeshine (Boeckh, 1975:
no information
whether referring to all cat species or domestic cats only).
Rodents: flying squirrels,
for instance
Petaurista
petaurista, in Malaysia: yellow to orange (F. Wiens, pers.
comm.).
(Barrett, 1984: white or lemon-yellow reflexion, somewhat
similar to N.
coucang)
Fruit-bats (e. g. Pteropus):
tapetum
present, no information about colour of eyeshine (Martin 1990)
Birds: Long-tailed night-jars
(Caprimulgus
macrurus) have a colour of eye-shine similar to that of N.
coucang,
but usually only one eye is visible, and the animals are found
sitting
in characteristic places, on lookouts such as masts (F. Wiens,
pers. comm.)
Sympatric species in
Africa with
reflecting eye background:
Galagos (Galagoides
allenii,Galagoides
demidoff, Euoticus elegantulus): orange to whitish
eyeshine?
(Colour seen in G. moholi in Bochum and on photos of
several species
in Rowe, 1996; Martin 1990); galagos can be identified by their
characteristic
calls (Charles-Dominique; 1971; Bearder et al., 1995, Ambrose,
Bearder,
1995). Usual time, frequency of calling?
Sympatric species in
the
different regions of distribution area which can be mistaken for
slender
lorises because of size, appearance, similar vocalizations or
movements?
(Viverrids, martens, arboreal cats, other arboreal species?
Birds? Similarity
of lorises with small owls?) See under references: literature
about sympatric
species.
Behavioural aspects:
Visible
behaviour:
movements helpful for identification of species; protective
behaviour (flight, camouflage)
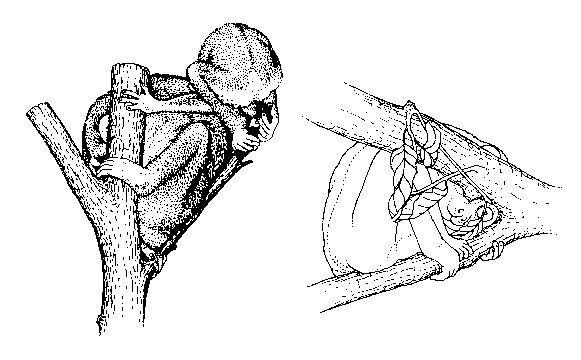
Hiding of the face or hiding
are
typical behaviours in lorises when disturbed (see figure).
The
manner in which slender lorises glance at and look away from the
light
is helpful for identification in connection with eye reflektion
(B. Meier,
unpublished; Nekaris 1997). According to Petter and Hladik
(1970), Loris
eye reflection in the wild was seldom visible for more than a
second whereas
nocturnal Malagasy lemurs looked towards the light for rather
long periods.
Barrett (1984) mentioned that observation with red light does
not cause
the eye-blinking pattern otherwise characteristic for lorises.
In the captive
Loris
tardigradus nordicus at Ruhr-University, hiding of the
face / turning
the face away from an observer is regularly shown as a sign of
stress when
shy animals are approached by a human. In spite of their usual
curiosity,
shy animals may freeze for longer periods in a posture in which
they cannot
see the approaching human. This is probably a camouflage
behaviour (hiding
of the conspicuous facial markings). Kingdon (1997) confirms for
Arctocebus
c. calabarensis in the wild that normal census methods
with use of
torchlights are unsuccessful because the animals hide their
heads and eyes
at the least disturbance. Slender lorises habituated to
observation may
look into a torchlight for somewhat longer periods when curious,
but they
may also look aside as though they dislike the light shining
into their
eyes. In captivity, slender lorises are cautious when perceiving
something
unfamiliar, but they may become well habituated to a wide
variety of regularly-occurring
stimuli. Considerable difference in the reactions towards
stimuli have
therefore been observed. In the wild, too, different local
habituation
of animals, for instance to cars or people, might lead to
different survey
results; and adaptation of survey methods to local conditions
might improve
the results.
Freezing for camouflage
apparently
is a behaviour typical for all Lorisinae / Loridae; it is common
in captive
Loris
and is also reported for Arctocebus and Perodicticus
in the
wild (Charles-Dominique, 1971). Inconspicuous, silent flight,
particularly
upwards, and hiding behind foliage or other cover are typical
protective
behaviour of the captive L. t. nordicus of
Ruhr-University when
being disturbed and are as well described for Loris from
the wild
(B. Meier, unpublished; Petter, Hladik 1970). According to
Charles-Dominique
(1977), pottos in the wild, noticing an observer in fairly close
proximity
(about 20 m) in all cases became immobile. When approach
continued, they
fled; when the observer remained immobile for 1-2 minutes, they
very slow
moved away, moving limbs one-by-one, immediate freezing at the
slightest
movement of the observer. This reaction in addition was
dependant on the
quality of environment: the animals fled rapidly if hiding in
the proximity
was possible; where environment provided no opportunity to
escape, for
instance in isolated trees, freezing was more frequent. In N.
coucang,
F. Wiens (pers. comm.) observed similar behaviour.
On the other hand, in Loris,
captive observation showed that cautious approach and
exploration of sources
of unfamiliar stimuli is a very characteristic behaviour,
apparently
vital for the animals´ safety (Schulze, Meier 1995). Balance
between
curiosity and caution was found to be dependant on individual
differences
and experience of animals and on the kind and size of stimuli:
small visible
objects were approached quicker and more readily than otherwise
equal large
objects (T. Milinski and S. Frank, unpublished), noise of
rattling of breaking
twigs clearly caused flight or hiding. During search for Loris
in
Sri Lanka with a head-lamp or car spot-light, the following
strategy worked
well: switching off the light at intervals and remaining
completely silent
and motionless for at least 1-2 minutes, lying motionless on the
ground
or car top, avoiding conspicuous eye movement or glintering of
teeth (captive
lorises show a fine perception of slightest movements). The
lorises then
apparently tried to find out where the origin of the disturbance
had vanished,
climbed an outlook or even approached the observer, and suddenly
switching
on the light again and quickly trying to scan the forest showed
their eye-shine
(B. Meier, unpublished). F. Wiens mentions for N. coucang
that animals
habituated to observation often did not look towards the
observer. In such
cases, waving with the torch-light often helped. (See also
above: red light).
MacKinnon and Phillips (1993) mention "pishing" (making
sibilant, squeaking
or rasping sounds) to make certain bird species approach and
investigate
the source of the sound.
Places / altitude where the
species
have been found in earlier studies:
Loris: occurrence
reported
higher in large trees (L. t. grandis in rainforest near
Kandy) to
low altidude in bushes; occasionally walks on the ground.
N. coucang: as in Loris
Arctocebus:
prefer
lower altitude; detection therefore less difficult than in
pottos. Prefer
substrate with small diameters, particularly vertical lianes
(Charles-Dominique,
1971).
In Perodicticus:
higher
in the trees, detection very difficult (Charles-Dominique,
1971).
Search
strategies:
During search for Loris
in
Sri Lanka in 1979 and 1980 (B. Meier, unpublished, duration: 12
weeks),
a pivoted spot-light on top of a Landrover was used to spot the
occurrence
of Loris eye-reflection at a speed of 10-15 km/h. To
test the method,
the same road was repeatedly searched on 10 different days, and
15 of 20
times lorises were seen in the same areas.
MacKinnon and Phillips (1993)
mention
that the speed of movement of an observer through the forest has
a considerable
influence on the result of surveys: an observer moving slowly in
order
not to miss anything will see lots of individuals but will be
likely to
miss the species which silently move away and hide when noticing
him. Quick,
silent moving or quiet sitting for a while may lead to
observation of animals
never seen otherwise. MacKinnon and Phillips recommend a
combination of
all three methods by turns for survey, with observers being
silent and
dressed in drab, never white clothes.
Activity peaks may also play
a role
in the probability to detect animals (MacKinnon and Phillips,
1993). For
lorises and pottos, studies indicate activity throughout the
night with
or without some increase of activity in the beginning and end of
the night
(Seitz 1967; Durrell 1949, quoted by Seitz; Baudenon 1949,
quoted by Seitz,
data from Ruhr-University). In the captive lorises at
Ruhr-University,
activity regularly decreased after consumption of larger amounts
of food.
Strong wind may make
detection and
observation easier because the animals climb down to lower
altitudes (Streicher,
pers. comm.).
Gill et al. (1997) mention
use of
portable thermal imaging for estimating deer population density
in forests
with varying visibility, "clearly revealing warm-bodied animals
even if
partly obscured by vegetation", animals in this study could be
detected
at so long distances that disturbance was minimized or
prevented; lorises,
however, are much smaller, usually living in a rather warm
environment
and keep at least their distal parts cool in order to save
energy; so it
seems doubtful whether infrared wave detection can also help to
detect
them
Charles-Dominique and Bearder
(1979)
used traps in addition to searching for rainforest lorisids and
caught
several pottos in an area in which a survey had been
unsuccessful. In the
course of a field study on slender lorises, a total of 1150
hours of trap
nights in the areas where animals were sighted, with sixteen
Tomahawk and
four Havahart live traps baited with honey -banana bait of
Tomahawk Trap
Company, ripe fruits and crickets, remained without any success,
animals
were then obtained by climbing trees and catching them by hand
(Kar Gupta
1995; Kar Gupta 2001: pers. comm). Detailed information about
mark-recapture
and other trapping for surveys is provided in Skalski and Robson
(1992)
and Sutherland (1996 a). In the paper on hand, no
recommendations how to
catch lorisids with traps are included in order to avoid use of
such information
for other than conservation purposes. Some captive slender
lorises at Ruhr-University
showed behavioural changes for 1-2 years after traumatic events
(catching,
transfer to unfamiliar environment), apparently trying to avoid
certain
stimuli reminding them of the stressful situation. In house
mice, individual
reaction to traps ("trap-proneness" and "trap-shyness") were
found to be
similar in parents and offspring and apparently independent of
experience
in traps (Crowcroft and Jeffers, 1961).
Charles-Dominique and Bearder
(1979)
tried to minimize traumatic effects of capture by regular
inspection of
traps during night to avoid animals being kept prisoner for a
longer time;
in animals caught with insufficient care, re-catching turned out
to be
very unlikely. Harcourt (1987) set traps (for Microcebus) for a
few hours
only and avoided to set them in rainy nights. Kenward (1987)
describes
a possibility to control traps by magnet-operated radio
transmitters (magnet
pulled off the transmitter when the trap is operated).
Sutherland (1996 b) mentions
hair
collecting facilities which may help to determine occurrence of
species
without traumatic capture. Hair tubes ( for smaller mammals) are
tubes
of a width slightly larger than the study species with
double-sided sticky
tape stuck to the inside; hair catchers are facilities with
wire-brush-like
structures animals are encouraged to squeeze through. Such
facilities,
baited ore just attached where animals are likely to pass, are
left in
the field for 1-2 weeks. For a lynx study, a "perfume"
containing pheromones
was developed for use with hair catchers; the animals rub their
head glands
on the scent bait, trying to deposit their own marks (Karge,
1999). Hair
catchers can be used in combination with a reference collection
or identification
key for hair of sympatric species or for DNA studies.
Vocalization:
In Loris, territorial whistling, chittering and
other
vocalization occur (Schulze, Meier 1995). According to Petter and
Hladik
(1970), humans can hear Loris whistling over a distance of 100 m.
Singh
et al. (2000) used territorial whistles for detection during a
survey.
In Zimmermann (1995 a, b), oscillograms and sonagrams of whistle
calls
of Loris, Nycticebus coucang, N. pygmaeus
and Perodicticus
potto are published.
Further
information
about vocalization in our behaviour database.
Voice recognition? Sympatric
species
with vocalizations which can be mistaken for loris / potto
vocalization?
The ornithologist Bill Evans, Cornell University, Ithaca,
developed a voice
recognition software which compares vocalization with a
"library" of sonagrams
of species; it identifies calls more reliably than even human
observers
and can record the presence of species in the field without
observers being
present ("Nickerchen ...", 1995)
Differences in vocalization
within
genera, species / subspecies differences? Vocalizations of large
grey and
small brown or reddish Loris forms from Sri Lanka, kept
at Ruhr-University,
sounded similar; there was no evidence for vocalization
differences helpful
for distinguishing slender loris forms as described in galagos
(Bearder
et al., 1995, Ambrose, Bearder, 1995).
Bird-watchers use playback of
tape-recorded
vocalization to cause a reaction; MacKinnon and Phillips (1993),
however,
mention that too frequent use particularly of tape recordings of
territorial
calls may disturb territorial behaviour and reduce the natural
alarm responseds
of the animals. In captive slender lorises, aggressive behaviour
between
formerly peaceful mates occurred as a consequence of territorial
whistles
by animals in neighbouring cages.
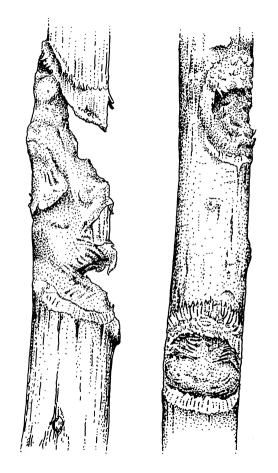
Traces
In N. coucang and N.
pygmaeus,
gnawing of branches may leave visible traces. Gnaw marks found
in a tree
species in Vietnam at an altitude of 8 m after observation of
pygmy lorises
had an average diameter of 2.5 cm and were encrusted by dried
exudates,
suggesting that lorises had supplemented their diet with gum or
sap (no
feeding behaviour was observed). Gnaw marks produced in
captivity might
be a sign of boredom (Tan 1994; Schweigert, pers. comm.;
Streicher, pers.
comm.; gum-feeding in N. coucang reported by Barrett
1984).
Gauge marks of captive Nycticebus coucang (redrawn
from a photo of branches in the collection of K.-H.
Schweigert)
Loris faeces
Faeces, smell:
Loris
faeces are small, firm and fall down where the animals
are just active.
Slender lorises do not fecally mark branches, and in the
captive colony
of Ruhr-University there were no places where
conspicuous quantities of
faeces were deposited. Territories are marked with small
quantities of
urine deposited on branches; such markings are optically
inconspicuous,
but easily watersoluble, some urine will probably be
washed down by rain.
During normal emission of urine, some drops to the
ground. Detection of
lorises, their faeces or urine with trained dogs might
be a possibility
to improve survey results. Zwickel (1980) gives a review
of possibilities
to use dogs in wildlife management and provides
addresses where to obtain
further unpublished information. He mentions that dogs
may be useful in
studies of marking behaviour, activity sites and refuge
behaviour, that
collection with dogs gives almost unbiased data on food
habits of some
species and less sex and age biased results than for
instance catching
of animals with traps. Particularly in animals located
higher in trees
behind foliage, dogs might be a valuable help.
Offering of food mixed with
indigestible
markers to animals might help to get information about range
sizes. Kenward
(1987) proposes coloured fibres; in tests with otters (Schulze,
unpublished),
small parts of coloured feathers and different small seeds
worked well.
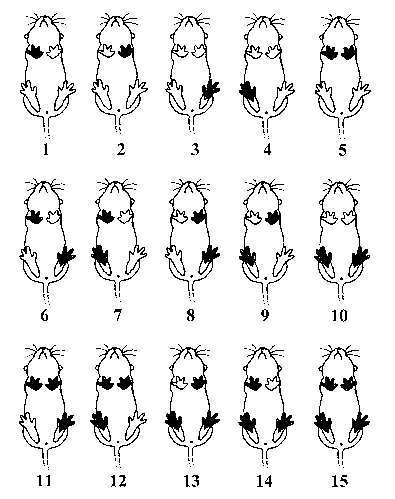
Individual
recognition,
marking of animals:
Numerical
marking up to 15 by marking
different limbs. Based on Renner, 1996. Dying of
the fur or cutting ring-shaped
gaps into the fur around the limbs might work in
lorises (not yet tested).
Nocturnal
conditions (changed colours, general vision) in
connection with the different
altitude of animals in the trees make an individual
identification and
estimation of size difficult (F. Wiens, pers. comm.).
Catching and marking
wild animals may help to determine population density
without counting
individuals twice. But marking requires consideration of
consequences for
the animals, including for instance: susceptibility to
traumatic effects
of capture (capture shock in certain species leading to
death by chilling
or predation, see Kenward 1987, Ginsberg et al., 1995).
Effects on study
results caused by capture, handling and / or marking,
danger of strangling
or getting stuck, increased detectability of marked
animals to predators,
local laws, visibility of marks from the distance, their
longevity and
the number of animals supposed to be distinguished must
be considered.
Ryder (1978) describes possible traumatic effects of
different kinds of
immobilizing and other drugs on wild animals and
discusses humane methods,
including the use of "biodegradable" material, decaying
after the period
for which it is needed, for marking to minimize the need
for recapture.
Handling of animals
caught for marking
and examination; use of drugs: see separate chapter, in
preparation; some
information can be found
in our first
aid chapter, the chapter about self-protection
when handling these species which produce a toxin,
and the chapter about
anesthesia
and
tranquilizers.
A variety of
numerical marking codes
are in use, independant from the kind of marks used
(examples: see figures
on the next page; the ear marking code shown
represents one of several
systems in use, it makes a maximum of two marks per
ear necessary). Combination
of such codes with markings of different colours may
allow individual marking
of large numbers of animals. Rice and Kalk (1996)
provide a review of standard
marking methods, including addresses where to obtain
necessary equipment.
See also Stonehouse 1978.
Non-invasive temporary
marking:
Rice
and Kalk (1996) mention dyes (Nyanzol), bleach, paint sticks and
balls
of paint shot with a gun, causing marks with a duration of
usually less
than a month. Longer-lasting marks might be achieved with
certain human
hair dye or hair-clipping in certain parts of the body. Markings
with Nyanzol-D,
a black hair dye which has been repeatedly used in animal
studies, were
visible for 7 months, until mold. The powder is mixed with
water, for fixing
this dye solution to the fur hydrogen peroxide solution is
needed. The
two solutions can be mixed together to facilitate the process.
(Chihuahuan
desert lab manual: Prairie dogs website). Megna (2001) for
instance remommends
a mix of 12 cc water, 12 cc over-the-counter hydrogen peroxide,
48 cc rubbing
alcohol, and 1 heaping tablespoon of dye, stirred well, allowed
to sit
a while, stirred again and applied with a sponge-type
paint/craft brush.
More information see Laboratory Primate Newsletter 40 (3), July
2001: Marking
Monkeys - Nyazol D. The former dye factory „Nyanza Chemical
Company“ was
closed in 1978; but Nyanzol might be available from Albinal
Dyestuff Intl.,
www.albanildye.com
(Giles 2000, F. Loureiro, pers. comm.). In animals with dark fur
bleached
marks may be better visible. Day, Schemnitz and Taber (1980)
provide further
information about different dyes and possibilities to apply them
to animals,
for instance with a paint pistol by the Nelson Paint Company,
operated
by CO2, which allows dying of animals
with paint pellets
at a distance of 15 m. They mention that for instance picric
acid applied
on rabbits was still well visible after seven months. F. Wiens
(1995, unpublished)
noted that on slow lorises dark colour spots were well visible
from some
distance; light marks were less well visible.
Permanent, invasive
visible marking:
Tattoos are usually
not well
recognizable from the distance even if rather hairless parts are
tattoed,
and in some species tattoos are less permanent than in others
(Rice and
Kalk, 1996). Tattoing on the plantar side of feet or medial
surface of
the upper thigh has worked well in some lorises, but in other
individuals,
tattoos became unreadable or vanished. There is some evidence
that too
dense tattooing in Loris may lead to skin necroses and
subsequent
vanishing of the tattoos (observation at Ruhr-University). In
small ears,
ear tattoos may lead to extensive damage and atrophy of the ear
(Rice and
Kalk, 1996).
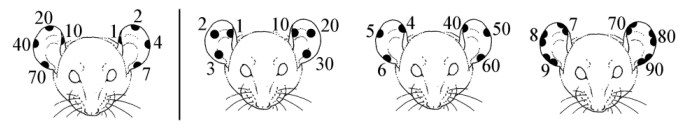
Ear-notching (cutting
notches
into the ear pinnae or using a special plier-punch) is commonly
used in
laboratory and zoo animals, but it is an amputation and not
allowed everywhere.
Ryder (1978) regards this marking method as not humane. Rice and
Kalk (1996)
mention excessive bleeding after ear-notching adult individuals.
In slender
lorises, blood-vessels leading along the ear rims probably play
a role
in the regulation of body temperature (see figure).
Laboratory animal
marking by ear
notching. Left: sites for numerical marking of the ear rim
up to number
100. Based on Rice and Kalk, 1996. Example: 68 = 1 + 7 +
20 + 40. Right:
marking method recommended by Renner, 1996. But see right
figure for possible
disadvantages of such an invasive marking method
Enlarged
bloodvessels in a Loris
ear during high ambient temperatures
Ear tags
Ear tags are commercially
available
in numerous sizes, different colours or with numbers. Tagging
means piercing
of the ear pinna; plastic tags seem to be less likely to cause
infections.
Size and thickness of the ear, location of stiffer cartilage or
large blood
vessels ought to be considered when chosing place and weight of
tags (see
also Rice and Kalk, 1996). Ear tags are well visible, but they
may be torn
off or get lost.
Freeze-marking
is a permanent, well-visible
marking
method which has been used for animals of the size from neonatal
mice to
elephants; it is regarded as painless because the tissue is
cooled down
to numbness and nerves are believed to be inactivated for
several weeks
by freezing. But the method still needs development, the final
well-visible
marks appear with delay, and good results depend on correct
marking: after
cooling of the skin to such a degree that melanocytes in the
hair follicles
are permanently destroyed, the superficial epidermis and hair is
shed within
weeks, and the new hair growing after 1-3 months is white.
Excessive freezing
causes scars instead, insufficient freezing causes incomplete
marks. Freeze-marking
can for instance be done with a thick copper tool cooled down in
a mixture
of dry ice and 95% alcohol (Rice and Kalk, 1996).
Attaching of collars, leg
bands,
radio transmitters and other tags:
Kenward (1987) mentions the
following
possible consequences of radio tag attachment besides possibly
traumatic
effects of capture, handling and presence of tags: temporary
increase in
grooming on costs of other behaviour, desertion of offspring by
tagged
parents or parents rejecting tagged offspring, skin lesions or
reduced
insulation and increased energy expenditure by damaged fur /
feathers because
of badly-fitting tags, reduction of agility in low weight
individuals.
Kenward recommends initial longerlasting tests with captive
individuals
of the species in question, when possible, before attaching tags
to wild
individuals. Broad collars are well visible, but need regular
control to
ensure that they do not cause health problems (Rice and Kalk,
1996). In
lesser slow lorises, collars might cause problems because of
seasonal weight
changes which may also alter neck circumference considerably (U.
Streicher,
Endangered Primate Rescue Center). Loss of weight in slow
lorises might
lead to loose collars increasing the danger that twigs stick
fast between
collar and neck; in fact broken twig remnants were found under a
collar
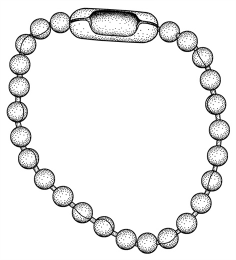
(F. Wiens, pers. comm.). Lightweight stainless metal bead chains
(see figure)
have been successfully used as collars in small mammals (E.
Curio, Ruhr-University
Bochum, pers. comm.; Luft, Curio 2001); such chains can for
instance be
obtained from PÖSAMO chain
factory
Pötz & Sand, Frohnstr. 44, 40789 Monheim, Germany, or
via
a czech PÖSAMO
factory. Luft and Curio successfully used these chains for
marking bats
and for attaching radio transmitters to them because they are
lightweight
(less than 0.7 % of body weight of the bats marked) and have no
sharp edges;
no dirt can stay under the rotating balls, causing skin
problems, and their
length is easily adjustable. Luft and Curio closed the chains
with stainless
steel locks with four-digits numbers laser-engraved on them for
individual
marking. The locks can be secured against loss by filling them
with some
glue; Luft and Curio closed them and in addition reduced
abrasion of fur
through the lock by covering it with soft silicon tubing (which
was semi-transparent
to allow reading of the numbers on the locks). Marking of
lorises with
such chains around the wrists? Hidden under fur, such chain
collars do
not allow identification without handling; individual
identification from
the distance might be possible by attaching short coloured
tassels to the
chain (see figures below), but this method has not yet been
tested.
In Loris, coloured
plastic
pigeon leg bands with an inner diameter of 8-9 mm, fixed around
the wrists,
turned out to be well visible even from some distance. The type
of rings
used (flat elastic tape rolled up) allows spreading of the rings
and attaching
them to the animals cautiously with the help of a second person.
Captive
animals carried such bands for years without any problems; only
in two
cases, lorises managed to get rid of their ring. In pygmy slow
lorises
at the Endangered Primate Rescue Center, Vietnam, no problems
caused by
the rings were observed, but in animals living in groups some
arm rings
were damaged by chewing or lost. Chewing was not observed, so it
is unknown
whether the animals themselves or conspecifics removed the rings
(U. Streicher,
unpublished). For slow lorises, at least the rather small pigeon
rings
cannot be recommended even though they are elastic and widen
when spread;
when such rings were tested on several confiscated slow lorises
in Indonesia,
one developed problems (swollen hand; after removal of the ring
a wound
was detected under it); the rings then were all removed (Femke
den Haas,
pers. comm.). F. Wiens (pers. comm.) made attempts with arm and
leg bands
on two wild slow lorises; both animals somehow managed to get
rid of the
ring.
Plastic bird rings might be
secured
by agglutination with acetone (B. Meier, pers. comm.)
Recognition marking with collars and bracelets ..
Stainless steel bead chain used for marking
fruit bats.
Redrawn from a photo by E. Curio, Philippine
Endemic Species Conservation Project. This type of
chain is lightweight,
can be well adapted in length and no dirt can accumulate
under it.
Coloured
bird ring
around the wrist of a pygmy loris. Photo: Ulrike
Streicher, Endangered
Primate
Rescue Center. This type of ring is too narrow for
use in larger
slow lorises, and under the broad ring dirt may
accumulate and cause
skin problems if it´s not loosely fitting. Loose
bracelets on the
other hand may increase the danger of getting caught in
some cleft. (In
captive slender lorises, however, this kind of bracelets
was used for many
years without causing problems)
Some more ideas for recognition marking with coloured
tassels
..
Tassels made of coloured fibers, attached to a bead chain
with loops. Photo
taken in daylight. Three tassels, left: material for
production of fly-fishing
lures; middle, straigt fibers: dyed artificial hair from a
barber´s
shop. Right: two tassels made of stripes of metal-like
reflecting plastic
foil. The tassel bases were made as slender as possible to
minimize the
risk that the animal gets hooked in any cleft. This
type of marking has not yet been practically tested.
..
The same tassels as in
the photo left,
seen with infrared night vision equipment (Sony Handycam
digital video
camera with night shot function). Colour differences are
invisible, reflecting
properties in infrared light apparently differ from
reflexion in daylight.
But differently reflecting tassels of varying structure
and lenght should
allow development of marks distinguishable with infrared
night vision equipment.
Photos: H. Schulze
..
Tassel made with stainless steel loop and Stabilit
epoxy glue.
..
Idea how to insert a piece of biodegradable material in a
bead chain: fine
leather shoe string attached in two bead chain locks with
knots. The string
may be further attached by filling the locks with glue.
Information about
the time after which biodegradable materials fall off
under different climatic
conditions is still lacking.
Use of "biodegradable"
material,
decaying after the period for which it is needed, as proposed by
Ryder
(1978, see also above), would certainly be a good idea in field
studies,
but at present information about practical solutions is lacking.
A recognition
mark caused to fall of by a radio signal would allow removal
after finishing
a study, minimizing the risk for the animal (see Kenward, 1987)
Light-emitting markers
may
allow easier retrieval of marked animals in darkness. (Risk that
such markings
also help predators to locate the animal?). Beta lights are
fluorescent
markers which may operate 10-20 years and are visible at 400 m.
They consist
of sealed glass capsules internally coated with phosphorus and
filled which
tritium gas which emits low-energy beta particles causing the
phosphorus
to glow. They are available in different sizes and colours, for
instance
from Saunders-Roe Development Ltd., Millington Road, Haynes,
Middlesex
UB3 4NB, U. K.; beta light in connection with radio transmitters
can be
obtained from Biotrack, Stoborough Croft, Grange Road,
Stoborough, Wareham,
Dorset BH20 5AJ, U. K., phone: x44-1929 552992, e-mail:
brian@biotrack.demon.co.uk
or sean@biotrack.demon.co.uk. For use of Beta light a permission
from radiation
safety authorities such as the Nuclear Regulatory Commission in
Washington
is necessary (B. Meier, pers. comm.; Lehner 1979 and 1996;
Kenward 1987;
MacDonald, 1978; Wolcott, 1980, Day, Schemnitz and Taber, 1980).
Lehner
(1996) mentions other methods for luminescent marking. Buchler
(1976) described
production and use of a chemiluminescent tag weighing less than
0.5 g,
filled with liquid from CYALUME chemical emergency light sticks,
which
gave light for several hours after mixing of the chemicals.
Lemen and Freeman
(1985) used a pigment powder, fluorescent under ultraviolet
light, with
low toxicity: small mammals were caught in traps, their fur
saturated with
the fine powder. After being released, the animals left traces
of the powder
during first night which persisted until next heavy rain.
So far, too little is known
about
the risks caused by attaching collars, bracelets or other marks
to wild
animalls over a longer period.
Hair clipping on one or
several
limbs is probably the least dangerous method for producing
recognition
marks visible from the distance
Survey
/
census methods:
Sampling is done for
determination
of species, of numbers of individuals in a given area or
population, population
dynamics and answers to many ecological questions. For
development of a
sampling program, one essential step is the comparison and
evaluation of
potential sampling techniques. Usually this is done by randomly
locating
sites in a habitat and collecting one sample unit by each
sampling technique
being considered; care should be taken that one technique does
not interfere
or bias another one (Pedigo, 1994; Buntin, 1994). Changes of
habitat quality
or behaviour of the species, for instance weather conditions,
annualperiodic
differences in density of foliage, changes of distribution /
abundance
of food, migration, occurrence of hibernation / torpor periods,
increased
or decreased caution in relation to breeding periods might
influence results.
Standardized methods for
determination
of population density in general and of average population
density in a
larger area with uneven distribution or differences in
detectability of
speciemens: see literature about biological and geographic
survey methods
below, under references
Some terms concerning
population
survey methods:
Sample / sample
population:
part of a larger population examined
Sampling: basic
quantitative
collection of population data, for instance concerning density,
dispersion,
age structure or other properties of the population, from a part
of a population,
subsequently generalising from the sample to the whole
population. Samples
must be representative of the whole, otherwise generalisation to
the whole
will produce biased results (based on Greenwood, 1996, changed.
Random sampling: n
possible
samples with the same probability for each sample to be selected
Count / direct count:
counting
individuals for instance in a study population / area / herd
Census / true census:
counting
all individuals in a population (Pedigo, 1994)
Survey: usually an
organized
program with detailed, established protocols for monitoring
animals (Higley,
Peterson 1994)
Monitoring: studying
the
development of populations or their environment over a period of
time,
using the same techniques of monitoring each time or, if the
decision is
made to change techniques, with a time of overlap in which both
methods
are used to determine the relative efficiency of both for
calibration of
data (adapted from Greenwood, 1996); broad sense, often only
qualitative
sampling used to trigger more precise quantitative sampling
programs, for
instance in seasonally occurring species (Higley, L. G.;
Peterson, R. K.
D., 1994). (Greenwood, 1996, used the term "surveillance" for
observation
of changes over time).
Population index:
measurement
related to the actual total number of animals (ideally the ratio
of index
to number should be constant: I/N = K; I
=
population index, N = number of individuals, K =
index ratio,
K
may be known or unknown). Examples of indices: number of animals
seen while
standing in a place during a certain period or while walking a
certain
distance or number of calls heard in a standard period. Indices
can be
compared to other indices (for instance of different places or
times).
Judgement of comparability, however, depends on knowledge of the
examined
species, because detectability, frequency of calls or other
counted events
may vary. Standardisation, for instance similar weather
conditions,the
same time of day or season and similar methods for data
collection or multiple
randomized observations are necessary. (Based on Greenwood,
1996, changed).
Some terms used for
describing
properties of populations: (based on Pedigo, 1993)
Absolute population
density:
total count or estimate of all individuals in a given area
(Pedigo, 1994).
Absolute population
estimates:
actual numbers of individuals on a given surface-area unit
(Pedigo, 1994).
Estimates of the total
population:
counting all individuals of a representative unit of the
population (for
instance in a certain area) (Pedigo, 1994)
Basic population estimates:
number of individuals per standard unit of habitable space. The
standard
unit for a species / population, for instance ft² of branch
surface,
may be established formally by a group of researchers or
informally through
traditional practice (Pedigo, 1994).
Relative population
estimates:
no direct relationship to land surface area, but referenced to
the sampling
technique used (Pedigo, 1994)
Density: an expression
of
abundance of a species / population in an area
Dispersion: the spatial
distribution of individuals of a species / population
Natality: birth rate
Mortality: death rate
Age structure: relative
proportions of individuals in different age classes
Population growth
(population
growth form): shape of population growth curve
Population density of
Lorisinae
/ Loridae: methods used in earlier studies and general
considerations concerning
lorises or pottos or their habitatsSingh et al. (1999)
recommend "fixed
point line transects" (Burnham et al. 1980) as the best method
because,
although males on occasions moved as far as 00 m durinng a
single night´s
activity, females seldom moved more than a few metresEstimation
of number
of animals remaining undetected, dependant on density of
vegetation,
altitude of trees, preferred altitude of stay? Charles-Dominique
and Bearder
(1979) recommend to note the distance of detected animals from
survey pathways
for every type of vegetation (with use of a surveyor´s tape or
calibrated
string, when necessary, because distances in the rainforest are
easily
overestimated) and to consider only the strips on either side of
the path
for population density estimates in which no reduction of the
number of
animals because of distance from the path was found. At Makokou,
Gabon,
the following distances for fairly good detection of animals
were determined:
Undergrowth of primary forest: 30m. Canopy of primary forest:
30m. Flooded
primary forest: 20m. Secondary forest and tree-fall zones: 20m.
River banks
surveyed from a canoe: 10 m
Danger of wrong population
density
estimates because of patchy or uneven distribution of animals?
A 12-week search in Sri Lanka
in
1979 and 1980 (Meier and Nieschalk, unpublished) showed that in
most examined
areas lorises appeared to be absent even when the habitat
appeared to be
suitable, whereas in several pockets of 50-100 km² high
population
densities were found for no evident reason. Data of densities
only from
locations which are chosen for surveys because of occurence of
lorises
at higher densities might therefore cause misleading population
density
estimates and underestimation of threat. Standardized methods
for determination
of population density in general and of average population
density in a
larger area with uneven distribution? See literature below.
Danger of wrong estimation of
population
density if survey pathways are leading along types of habitat
different
from average forest cover? For the Bornean rainforest, MacKinnon
and Phillips
(1993) mention that forest edges with light falling in sideways,
for instance
along roads, are very rich feeding zones with a rather dense
bird population,
better observation possibilities and particularly inhabited by
"forest-edge
species". Literature reports (Petter, Hladik 1970, Johnson 1984)
indicate
that slender lorises were detected close to roads or in forest
rims close
to water, but it is not quite clear whether this was due to use
of roads
or boats for searching or due to a preference of forest rims by
the animals.
Is the distribution constant
or
does it change, for instance with seasons? Petter and Hladik
(1970) mention
L.
t. nordicus which during nine nights in December / January
were found
close to a lake, whereas in May and October several animals were
detected
close to a road a few hundred meters further east, more distant
from the
shore of the lake. They concluded that in slender lorises the
location
of range / teritory may vary in the course of the year. So far,
no reports
about longer or regular migrations have been found. Slender and
slow lorises
may walk over the ground to reach distant trees (Petter, Hladik
1970; F.
Wiens and A. Zitzmann, pers. comm.).
Some
literature
concerning collection of habitat data:
Basic methods for habitat
analysis
and description are described by Gysel and Lyon (1980) and Jones
and Reynolds
(1996). Korschgen (1980) provides a review of procedures for
food-habits
analyses, such as sampling and analysis of stomach contents and
faeces;
further information about methods for analysis of faeces are for
instance
provided in Dickmann and Huang (1988). Hutchins (1994) gives a
review of
techniques used for quantitative sampling of arthropods. Nagy
and Haufler
(1980) describe analysis methods for determination of forage
nutritive
quality.
Some
literature
references: planning conservation projects
Caldecott, J. O., 1996: Designing conservation projects:
People
and Biodiversity in Endangered Tropical Environments. Cambridge
University
Press, Cambridge. ISBN/ISSN 0-521-47328-4 (Hardcover).
Margoluis, R.; Salafsky, N.; Balla, A. (Illustrator),
1998: Measures
of Success : Designing, Managing, and Monitoring Conservation and
Development
Projects. Island Press. ISBN: 1559636122 (Paperback, 363
pages)
Some
literature
references: field methods, biological and geographic survey
methods
Feinsinger, P., 2001: Designing Field Studies for
Biodiversity
Conservation: The Nature Conservancy. Island Press. ISBN:
1559638788 (Paperback:
224 pages)
Barnett, A., 1995: Primates: Expedition Field Techniques.
By
Expedition Advisory Centre, Royal Geographical Society, London.
Boitai, L.; Fuller, T. K., 2000: Research techniques in
animal
ecology: controversies and consequences. Columbia University
Press, New
York.
ISBN: 0-231-1134-4 or ISBN 0-231-1134-2.
Giles, R. H. (ed.),
1972:
Wildlife management techniques. 3rded.,
rev. The Wildlife Society, Washington, D. C. ISBN:
0-933564-08-2.
Giles,
R.
H. (ed.), 1969: Wildlife management techniques. The
Wildlife Society,
Washington, D. C.
Hayek, Lee-Ann C.; Buzas,
M.
A., 1997: Surveying natural populations. Columbia
University Press,
New York. Hardback. ISBN: 0-231-10240-2.
= Hayek, Lee-Ann C.;
Buzas,
M. A., 1998: Surveying natural populations. Columbia
University Press,
New York. Paperback.
Content:
statistical
principles for quantitative population surveys.
Hill and Clayton, 1985: Wildlife after dark: a review of
nocturnal
observation techniques. Occasional paper no. 17, James Ford Bell
museum
of Natural History, University of Minnesota, Minneapolis. 23 pp.
(Quoted
in Lehner 1996)
Kenward, R., 1987:
Wildlife
radio tagging. Equipment, field techniques and data analysis.
Academic
Press, London.
Lehner, Philip N.,
1996:
Handbook of ethological methods, 2. ed. Cambridge Univ. Press,
Cambridge.
672 pp. ISBN 0-521-55405-5
Lehner,
Philip
N., 1979: Handbook of ethological methods. Garland STPM
Press, New
York, London. ISBN: 0-8240-7024-0.
Sutherland, William J.,
1996
a: Ecological census techniques: a survey. Cambridge University
Press,
Cambridge. ISBN: 0-521-47244-x.
Content:
survey methods;
methods for estimation of population size in plants and
animals; methods
for measuring environmental variables such as weather, water
chemistry
and soil composition.
Skalski, John R.; Robson,
D.
S., 1992: Techniques for wildlife investigations. Design
and analysis
of capture data. Academic Press, San Diego (u. a.). ISBN:
0-12-647675-6
Content:
capture
- marking - recapture data: evaluation, incorporation into
population investigations;
quantification in changes of animal abundance, assessment of
environmental
impact.
Ritchie, William; Wood,
M.; Wright,
R.; Tait, D., 1988: Surveying and mapping for field
scientists. Addison
Wesley Longman ISBN: 0-582-30086-x.
Content:
geographic
surveying techniques; mapping.
Ritchie, William; Tait, D.
A.;
Wood, M.; Wright, R., 1977: Mapping for field scientists.
A problem.solving
approach. A. S. Barnes and Company, South Brunswick and New
York. ISBN:
0-498-02036-3.
Content:
ground,
aerial, photogrammetric geographic surveying; reading maps,
mapping: comprehensive
practical introduction for field scientists with no surveying
experience.
References
quoted
in the text:
Ambrose, L.; Bearder, S.
K.,
1995: Identifying primate species from their calls. Poster
abstract, International
Conference on the Biology and Conservation of Prosimians,
Chester, 13-16
September 1995.
in press? in: Folia
Primatologica
69
(Suppl. 1) (= Proceedings of the international conference on the
biology
and conservation of prosimians, Chester, 13-17 November 1995).
Barrett, E., 1984: The
ecology
of some nocturnal, arboreal mammals in the rainforests of
peninsular Malaysia.
Ph.D. Thesis, Cambridge University
Baudenon, P., 1949:
Contribution
à la connaissance du Potto de Bosman dans le Togo-Sud. Mammalia
13: 76-99.
Bearder, S. K.; Honess, P.
E.;
Ambrose, L., 1995: Species diversity among galagos with
special reference
to mate recognition. Pp. 331-352 in: Alterman, L.; Doyle, G. A.;
Izard,
M. K. (eds): Creatures of the Dark. The nocturnal prosimians.
New York,
Plenum Press.
Berry, R. J., 1970:
Covert
and overt variation, as exemplified by British mouse
populations. Symp.
zool. Soc. Lond. 26: 3-26.
Boeck, J., 1975:
Nervensysteme
ud Sinnesorgane der Tiere. Herder Verlag, Freiburg im Breisgau.
Buchler, E. R., 1976:
Chemiluminescent
tag for tracking bats and other small nocturnal animals. Journal
of mammalogy
57:
173-176.
Buntin, G. D., 1994:
Developing
a primary sampling program. Pp. 99-115 in: Handbook of sampling
methods
for arthropods in agriculture. Pedigo, L. P.; Buntin, D. G.
(eds.), CRC
Press, Boca Raton.
Burnham, K. P.; Anderson, D. R.; Laake, J. L., 1980:
Estimation
of density from line-transect sampling of biological populations.
Wildlife
monographs 72: 1-202.
Charles-Dominique, P.,
1971:
Eco-ethologie des prosimiens du Gabon. Biol. Gabonica 7:
121-228.
Charles-Dominique, P.;
Bearder,
S. K., 1979: Field studies of lorisid behavior:
methodological aspects.
Pp. 567-629 in: Doyle, G. A.; Martin, R. D.: The Study of
Prosimian Behavior.
Academic Press, New York.
Chihuahuan desert lab
manual:
Prairie dogs website – Project 3. Study proposal / field
protocol:
Reintroduction and monitoring of prairie dog populations: a
study proposal.
http://www.nps.gov/cave/cdl/p3protocol.pdf. Seen on 29 June
2001.
Crowcroft, W. P.; Jeffers,
J.
N. R., 1961: Variability in the behaviour of wild house
mice (Mus
musculus L.) towards live traps. Proc. Zool. Soc. Lond. 137:
573-582.
Day, G. I.; Schemnitz, S.
D.;
Taber, R. D., 1980: Capturing and marking wild animals.
Pp. 61-88 in:
Schemnitz, S. D. (ed.): Wildlife management techniques manual.
The Wildlife
Society, Washington, D. C.
Den Haas, Femke,
quality
controller animal care and paramedic, pers. comm., 2003:
information from
work at the Schmutzer Primate Center, Ragunan Zoo, Jakarta.
Dickman, C. R.; Huang, C.,
1988: The reliability of fecal analysis as a method for
determining the
diet of insectivorous mammals. Journal of Mammology 69
(1): 108-113.
ISSN: 0022-2372
Durrell, G. M., 1949:
The
angwantibo. Zoo life, London, 4: 29.
Giles, R. H., Jr.,
2000 :
The trevey. A total forest management plan and wildland
management decision
support system. Website:
http://fwie.fw.vt.edu/rhgiles/Trevey/Deer.htm.
Seen on 29 June 2001.
Gill, R. M. A.; Thomas, M.
L.;
Stocker, D., 1997: The use of portable thermal imaging for
estimating
deer population density in forest habitats. Journal of applied
ecology
34
(5): 1273-1286.
Ginsberg, J. R.;
Alexander, K.
A.; Creel, S.; Kat, P. W.; McNutt, J. W.; Mills, M. G. L.,
1995: Handling
and survivorship of African wild dog (Lycaon pictus) in
five ecosystems.
Conservation Biology 9 (3): 665-674.
Greenwood, J. J. D.,
1996:
Basic techniques. Pp. 11-110 in: Sutherland, William J.:
Ecological census
techniques: a survey. Cambridge University Press, Cambridge.
Gysel, L. W.; Lyon, J.,
1980:
Habitat analysis and evaluation. Pp. 305-326 in: Schemnitz, S.
D. (ed.):
Wildlife management techniques manual. The Wildlife Society,
Washington,
D. C.
Harcourt, C., 1987: Brief trap and retrap study of the
Brown
Mouse Lemur (Microcebus rufus). Folia Primatologica 49:
209-211
Higley, L. G.; Peterson,
R. K.
D., 1994: Initiating sampling programs. Pp. 119-136 in:
Handbook of
sampling methods for arthropods in agriculture. Pedigo, L. P.;
Buntin,
D. G. (eds.), CRC Press, Boca Raton.
Hladik, C. M.; Petter, J.
J.,
1970: Le loris tardigrade. Observations de terrains effectuées à
Ceylan. Science et Nature 101.
Hutchins, S. H., 1994:
Techniques
for sampling arthropods in integrated pest management. Pp. 73-97
in: Handbook
of sampling methods for arthropods in agriculture. Pedigo, L.
P.; Buntin,
D. G. (eds.), CRC Press, Boca Raton.
Johnson, J. M., 1984:
Diurnal
activities of the slender loris, Loris tardigradus, in the
Mundanthurai
Sanctuary, Tamil Nadu (India). Pp. 389-395 in: Current primate
researches,
M. L. Roonwal; S. M. Mohnot; N. S. Rathore (eds.), Jodhpur
Jones, J. C.; Reynolds, J.
D.,
1996: Environmental variables. Pp. 281-316 in: Sutherland,
William J.:
Ecological census techniques: a survey. Cambridge University
Press, Cambridge.
Kar Gupta, K., 1995: Slender loris, Loris tardigradus,
distribution and habitat use in Kalakad-Mundanthurai Tiger
Reserve, India.
Folia Primatologica 69 (suppl.): 401-402.
Kar-Gupta, K., in preparation: A study of distribution and
habitat
use by slender loris in Kalakad-Mundanthurai Tiger Reserve, India.
Accepted
with revision.
Karge, D., 1999:
Verräterische
Nagelkissen. Bild der Wissenschaft 10: 65.
Kenward, R., 1987:
Wildlife
radio tagging. Equipment, field techniques and data analysis.
Academic
Press, London.
Kingdon, J., 1997: The
Kingdon
field guide to African mammals. Academic Press, San Diego,
London. ISBN:
0-12-408355-2
Korschgen, L. J.,
1980: Procedures
for food-habits analyses. Pp. 113-127 in: Schemnitz, S. D.
(ed.): Wildlife
management techniques manual. The Wildlife Society, Washington,
D. C.
Lehner, Philip N.,
1979:
Handbook of ethological methods. Garland STPM Press, New York,
London.
ISBN: 0-8240-7024-0.
Lehner, Philip N.,
1996:
Handbook of ethological methods, 2nd edition. Cambridge
University Press,
Cambridge. ISBN 0-521-55405-5
Lemen, C. A.; Freeman, P.
W.,
1985: Tracking mammals with fluorescent pigments: a new
technique. Journal
of Mammalogy 66 (1): 134-136. ISSN: 0022-2372
Content:
dye-maring,
fluorescent pigments use in tracking; new technique, nocturnal
activity
Luft, S.; Curio, E., , 2002: An improved necklace for
marking
fruit bats. Ecotropica 8: 249-251..
MacDonald, 1978:
Radiotracking.
Some applications and limitations. Pp. 192-204 in: Animal
marking: recognition
marking of animals in research, B. Stonehouse (ed.), MacMillan,
London.
MacKinnon, J.; Phillips,
K.,
1993: The birds of Borneo, Sumatra, Java and Bali. Oxford
University Press,
Oxford. ISBN: 0-19-854035-3.
Martin, R. D., 1990:
Primate
origin and evolution. A phylogenetic reconstruction. Chapman and
Hall,
London
Megna, N. and others,
2001,
in Marking Monkeys - Nyazol D. Laboratory Primate Newsletter 40
(3), July 2001, online edition in
http://www.brown.edu/Research/Primate/lpn40-3.html#dye.
Nagorsen, David W.;
Peterson,
R. L., 1980: Mammal collector´s manual: A guide for
collecting,
documenting and preparing mammal specimens for scientific
research. Life
Sciences Miscellaneous Publications, Toronto: Royal Ontarion
Museum. ISBN:
0-88854-255-0.
Nagy, J. G.; Haufler, J.
B.,
1980: Wildlife nutrition. Pp. 129-142 in: Schemnitz, S. D.
(ed.): Wildlife
management techniques manual. The Wildlife Society, Washington,
D. C.
Nekaris, K. A.: Loris
internet homepage: http://artsci.wustl.edu/~kabiela
Nekaris, K. A., 1997:
A preliminary
survey of the slender loris, Loris tardigradus, in South
India.
Sixty-sixth Annual Meeting of the American Association of
Physical Anthropologists,
St. Louis, Missouri, USA, April 1-5, 1997. American Journal of
Physical
Anthropology, 0 (SUPPL. 24). 176-177. ISSN: 0002-9483
Pedigo, L. P., 1994:
Introduction
to sampling arthropod populations. Pp. 1-11 in: Handbook of
sampling methods
for arthropods in agriculture. Pedigo, L. P.; Buntin, D. G.
(eds.), CRC
Press, Boca Raton.
Petter, J. J.; Hladik, C.
M.,
1970: Observations sur le domaine vital et la densité de
population
de Loris tardigradus dans les forêts de Ceylan. Mammalia
34
(3): 394-409.
Renner, P., 1996:
Aufgaben
des Tierpflegers bei der Erstellung von transgenen Mäusen. Der
Tierschutzbeauftragte
3:
213-218.
Rice, C. G.; Kalk, P.,
1996:
Identification and marking techniques. Pp. 56-66 in: Kleimann,
D. G.; Allen,
M. E.; Thompson, K. V.; Lumpkin, S.; Harris, H. (eds.): Wild
mammals in
captivity, Principles and techniques. The University of Chicago
Press,
Chicago.
Rowe, N. (ed.), 1996:
The
pictorial guide of living primates. Pogonias Press, New York.
Ryder, R. D., 1978:
Postscript:
towards humane methods of identification. Pp. 229-234 in:
Stonehouse, B.
(ed.): Animal marking: recognition marking of animals in
research. University
Park Press, Baltimore. 224 pp.
Schulze, H.; Meier, B.,
1995:
Behavior of captive Loris tardigradus nordicus: a
qualitative description,
including some information about morphological bases of
behavior. Pp. 221-249
in: Creatures of the Dark, Alterman, L.; Doyle, G.; Izard, M. K.
(eds.),
Plenum Press, New York.
Seitz, E., 1967:
Untersuchungen
zur Aktivitätsrhythmik dunkelaktiver Halbaffen der Unterfamilie
Lorisinae.
Pp. 322-326 in: Neue Ergebnisse der Primatologie / Progress in
primatology,
1st Congress of the International Primatological Society.
Starck, D.; Schneider,
R.; Kuhn, H.-J. (eds.), Gustav Fischer Verlag, Stuttgart.
Singh, M.; Lindburg, D. G.; Udhayan, A.; Anand Kumar, M.;
Kumara,
H. N., 1999: Status survey of slender loris Loris
tardigradus lydekkerianus
in Dindigul, Tamil Nadu, India. Oryx 33 (1):
31-37.
ISSN: 0030-6053
Singh, M.; Anand Kumar, M.; Kumara, H. N.; Mohnot, S. M., 2000:
Distribution
and conservation of slender lorises (Loris tardigradus
lydekkerianus) in Southern Andhra Pradesh, South India.
International
Journal of Primatology 21 (4): 721-730.
Southern, N., 1955:
Nocturnal
animals. Scientific American 193 (4): 88-98.
Stonehouse, B. (ed.),
1978:
Animal marking: recognition marking of animals in research.
University
Park Press, Baltimore. 224 pp.
Streicher, U., 2000-2004: unpublished data from work as a
veterinarian
at the Endangered Primate Rescue Center, Vietnam, including data
from:
Streicher, U., thesis in
preparation.
Sutherland, William J.,
1996
b: Mammals. Pp. 260-280 in: Ecological census techniques: a
survey. Sutherland,
W. J. (ed.). Cambridge University Press, Cambridge.
Tan, C. L., 1994:
Survey
of Nycticebus pygmaeus in southern Vietnam. 15th
Congress of the
International Primatological Society: 136. (Abstract)
Wolcott, T. G., 1980:
Optical
and radio-optical techniques for tracking nocturnal animals. Pp.
333-338
in: A handbook on biotelemetry and radio tracking, Amlaner, C.
J.; Macdonald,
D. W. (eds.), Pergamon Press, Oxford.
Zimmermann, E., 1995a:
Acoustic
communication in nocturnal prosimians. Pp. 311-330 in: Creatures
of the
Dark, Alterman, L.; Doyle, G.; Izard, M. K. (eds.), Plenum
Press, New York.
Zimmermann, E., 1995b:
Loud
calls in nocturnal prosimians: structure, evolution and
ontogeny. Pp. 47-72
in: Current topics in primate vocal communication, Zimmermann et
al. (eds.),
Plenum Press, New York.
Zwickel, F. C., 1980:
Use
of dogs in wildlife biology. Pp. 531-536 in: Schemnitz, S. D.
(ed.): Wildlife
management techniques manual. The Wildlife Society, Washington,
D. C.
Some
literature
about species occurring in loris and potto habitats:
Vietnam:
Ziegler, T., 2001: Die
Amphibien
und Reptilien eines Tieflandfeuchtwald-Schutzgebietes in Vietnam
[Amphibians
and reptiles of a lowland rain forest reserve in Vietnam]. NTV
(Natur und
Tier Verlag GmbH), Münster. 344 pp., 367 colour figures, 77 maps
and
diagrams. Information about 100 species, based on surveys in the
region
"Ky An - Ke Go", southern North Vietnam. ISBN:
3-931587-54-1 (German)
India:
Bole, P. V.; Vaghani, Y.,
1986: Field Guide to the Common Trees of India. Oxford
University Press,
Bombay, Delhi, Calcutta, Madras.
Champion, H. G.; Seth, S.
K.,
1969: A Revised Survey of the Forests Types of India. The
Manager of Publication,
New Delhi.
Gurung, K. K.; Sing, R.,
previously 1998: Field guide to the mammals of the Indian
subcontinent:
where to watch mammals in India, Nepal, Bhutan, Bangladesh, Sri
Lanka and
Pakistan. Academic Press.
Content: 106
species
of larger mammals: identification in the field, habitat,
range, status.
Colour plates and figures of tracks.; review of national parks
and protected
areas.
Hutton, A. F., 1949:
Notes
on the snakes and mammals of the high wavy mountains, Madura
district,
South India. J. Bombay Nat. Hist. Soc. 48: 681-694
Prater, S. H. (ed.),
1965:
The book of Indian animals. Bombay Natural History Society.
First published
1948; reprinted with corrections 1980, several editions. Fourth
impression:
1993. ISBN: 0 19 562169 7.
Sahni, K. C., 1998:
The Book
of Indian Trees. Oxford University Press, Mumbai.
Srivastata, A., 1999:
Primates
of Northeast India. Megadiversity Press, Bikaner, India. ISBN:
81-87585-00-5:
Content:
Primates,
their habitats, food plants, conservation.
Whistler, H., 1963:
Popular
handbook of Indian birds. Oliver and Boyd, Edinburgh.
Sri Lanka:
Eisenberg, J. F.;
Lockhart, M.,
1972: An ecological reconnaissance of Wilpattu National Park,
Ceylon. Smithsonian
Contributions to Zoology 101: 1 - 119. Smithsonian Press,
Washington.
Eisenberg, J. F.; McKay,
G. M.,
1970: An annotated checklist of the recent mammals of Ceylon
with keys
to the species. Ceylon J. Sci. (Bio. Sci.) 8 (2): 69-99.
Henry, G. M., 1978: A
guide
to the birds of Ceylon. K. V. G. de Silva & Sons, Kandy, Sri
Lanka.
Phillips, W. W. A.,
1924:
A guide to the mammals of Ceylon. Ceylon Journal of Science,
Spolia Zeylanica
XIII (1): 261-283.
Phillips, W. W. A.,
1935:
Manual of the mammals of Ceylon. Colombo Museum, Ceylon.
Slow loris
distribution
areas:
Duckworth, J. W.; Timmins,
R.
J.; Thewlis, R. C. M.; Evans, T. D.; Anderson, G. Q. A.,
1994: Field
observations of mammals in Laos, 1992-1993. Natural History
Bulletin of
the Siam Society, 42 (2): 177-205. ISSN: 0080-9462
Davison, G. W. H.; Fook,
C. Y.,
1998: A Photographic Guide to Birds of Peninsular Malaysia and
Singapore.
Chelsea Green Pub Co; Paperback: 144 pages; ISBN: 0883590360.
Francis, C. M., 2001: A Photographic Guide to Mammals of
South-East
Asia: Including Thailand, Malaysia, Singapore, Myanmar, Laos,
Vietnam,
Cambodia, Java, Sumatra, Bali Borneo. Ralph Curtis Pub. Paperback:
127
pages. ISBN: 0883590522.
King, B. F.; Dickinson, E.
C.,
1975: A field guide to the birds of south-east Asia. William
Collins Sons
& Co Ltd, London.
Madoc, G. C., 1956: An
introduction
to Malayan birds. Published by The Malayan Nature Society.
Caxton Press
Ltd., Kuala Lumpur, Federation of Malaya.
Webster, M.; Fook, C. Y.,
1999: A Photographic Guide to Birds of Thailand. Paperback: 144
pages.
Chelsea Green Pub Co; ISBN: 0883590417







 The
manner in which slender lorises glance at and look away from the
light
is helpful for identification in connection with eye reflektion
(B. Meier,
unpublished; Nekaris 1997). According to Petter and Hladik
(1970), Loris
eye reflection in the wild was seldom visible for more than a
second whereas
nocturnal Malagasy lemurs looked towards the light for rather
long periods.
Barrett (1984) mentioned that observation with red light does
not cause
the eye-blinking pattern otherwise characteristic for lorises.
In the captive
Loris
tardigradus nordicus at Ruhr-University, hiding of the
face / turning
the face away from an observer is regularly shown as a sign of
stress when
shy animals are approached by a human. In spite of their usual
curiosity,
shy animals may freeze for longer periods in a posture in which
they cannot
see the approaching human. This is probably a camouflage
behaviour (hiding
of the conspicuous facial markings). Kingdon (1997) confirms for
Arctocebus
c. calabarensis in the wild that normal census methods
with use of
torchlights are unsuccessful because the animals hide their
heads and eyes
at the least disturbance. Slender lorises habituated to
observation may
look into a torchlight for somewhat longer periods when curious,
but they
may also look aside as though they dislike the light shining
into their
eyes. In captivity, slender lorises are cautious when perceiving
something
unfamiliar, but they may become well habituated to a wide
variety of regularly-occurring
stimuli. Considerable difference in the reactions towards
stimuli have
therefore been observed. In the wild, too, different local
habituation
of animals, for instance to cars or people, might lead to
different survey
results; and adaptation of survey methods to local conditions
might improve
the results.
The
manner in which slender lorises glance at and look away from the
light
is helpful for identification in connection with eye reflektion
(B. Meier,
unpublished; Nekaris 1997). According to Petter and Hladik
(1970), Loris
eye reflection in the wild was seldom visible for more than a
second whereas
nocturnal Malagasy lemurs looked towards the light for rather
long periods.
Barrett (1984) mentioned that observation with red light does
not cause
the eye-blinking pattern otherwise characteristic for lorises.
In the captive
Loris
tardigradus nordicus at Ruhr-University, hiding of the
face / turning
the face away from an observer is regularly shown as a sign of
stress when
shy animals are approached by a human. In spite of their usual
curiosity,
shy animals may freeze for longer periods in a posture in which
they cannot
see the approaching human. This is probably a camouflage
behaviour (hiding
of the conspicuous facial markings). Kingdon (1997) confirms for
Arctocebus
c. calabarensis in the wild that normal census methods
with use of
torchlights are unsuccessful because the animals hide their
heads and eyes
at the least disturbance. Slender lorises habituated to
observation may
look into a torchlight for somewhat longer periods when curious,
but they
may also look aside as though they dislike the light shining
into their
eyes. In captivity, slender lorises are cautious when perceiving
something
unfamiliar, but they may become well habituated to a wide
variety of regularly-occurring
stimuli. Considerable difference in the reactions towards
stimuli have
therefore been observed. In the wild, too, different local
habituation
of animals, for instance to cars or people, might lead to
different survey
results; and adaptation of survey methods to local conditions
might improve
the results.